- Auteur : D. CHARRON
- Ouvrage: Traité de Médecine (3ème édition), éd. Flammarion
- Date de publication: septembre 1996
Nous sommes conscients que cet article est relativement ancien; toutefois, il renferme des renseignements qui vous seront sans doute utiles; aussi l’avons-nous laissé en ligne. Par ailleurs, si vous disposiez d’articles plus récents sur le même sujet, n’hésitez pas à nous en faire part afin de demander leur autorisation à leurs auteurs pour une publication en ligne sur ce site
L’observation d’un contrôle genetique chez la souris de la prise ou du rejet de tumeurs greffees a conduit a la decouverte du CMH. Le systeme HLA, CMH de l’homme, est un ensemble de genes situes sur le bras court du chromosome 6, qui s’expriment principalement a la surface des cellules immunocompetentes sous la forme de glycoproteines transmembranaires. Les reactions allogeniques humorales et cellulaires survenant apres transfusion et transplantation ont permis de decouvrir la fonction physiologique principale de ces antigenes: le contr61e des reponses immunitaires contre les agents etrangers. Outre leur r61e essentiel en transplantation, leur interet medical est considerable car certains alleles du CMH conferent une susceptibilite ou une resistance genetique a de nombreuses pathologies, notamment auto-immures.
Le systeme HLA est le complexe genetique fonctionnel de l’homme le plus polymorphe, avec de nombreux loci exprimant pour la plupart de nombreux alleles (jusqu’à 8O). Cette diversite deja importante chez chaque individu est considerable au niveau de la population generale. Mis en evidence des 19S8 par des reactions serologiques d’ agglutination des globules blancs, puis par un test de microlymphocytotoxicite, le polymorphisme du systeme a ete disseque dans un premier temps grace aux approches classiques de la genetique, c’est-a-dire des etudes de segregation familiale et de repartition dans la population.
Deux classes principales d’antigenes ont ete decrites: les antigenes de classe I (HLA-A, B et C) presents sur la quasi-totalite des cellules de l’organisme et les antigenes HLA de classe II (HLA-DR, DQ et DP) restraints auxIymphocytes B, aux monocytes et macrophages. L’expression aberrante d’antigenes de classe 11 sur des cellules qui ne les expriment pas a l’etat physiologique est un des stigmates des etats auto-immune des rejets et des reactions du greffon contre l’hôte. Les molecules du systeme HLA de classe I et de classe 11 ont pour fonction principale de presenter les antigenes aux cellules T. Cette presentation est genetiquement << restreinte >> par le systeme HLA, et l’ensemble antigene peptide d’origine endogene ou exogene-molecule HLA (zone polymorphe) est reconnu par un recepteur specifique de la cellule T. 11 s’ensuit une activation T specifique. Les molecles HLA de classe I presentent un antigene peptidique originaire du cytosol de la cellule (proteine du soi ou proteine virale), avant d’etre transfere dans le reticulum endoplasmique. Les molecules HLA de classe 11 presentent des antigenes d’origine exogene localises dans les compartiments Iyso- endosomiaux apres leur endocytose. Les molecules accessoires (CD8 pour HLA classe 1, CD4 pour HLA classe 11) orientent la reponse immunitaire vers la cytotoxicite (cellules CTL CD8) ou la proliferation (cellules helper CD4), mais les mecanismes de reconnaissance et de restriction genetique sont identiques. La dichotomie classe I versus classe 11 a pu etre expliquee recemment, non par des differences structurales intrinseques des molecules HLA, mais par les voies intracellulaires -tutelles empruntent (exocytose pour les molecules de classe 1, endocytose pour les molecules de classe 11) et les partenaires antigeniques (peptides du soi et d’origine virale pour la premiere, peptides bacteriens et provenant de toxines pour la seconde) -tutelles rencontrent, chargent, transportent et vont presenter aux cellules T capables de reconnaitre leurs combinai sons specifiques .
Les reactions allogeniques au cours des greffes sont considerees comme des reponses antigeniques classiques ou l’allo-antigene devient lui- meme un antigene (seul ou associe a des peptides du soi) qui, reconnu comme << etranger >> sera responsable d’un rejet (CTL) ou d’une proliferation (GVH). Au cours des rejets, les Iymphocytes T cytotoxiques CD8 ami-classe I sont les effecteurs principaux, meme si des reponses cytotoxiques ami-classe 11 (CTL CD4) sont decelees. Dans la GVH, les reponses proliferatives CD4 anticlasse 11 predominent.
Les progres de la biochimie et de la biologie moleculaire permettent une vision precise tridimensionnelle du polymorphisme HLA et des interactions entre molecules HLA, peptides antigeniques et recepteurs T. L’obtention en 1987 de cristaux de molecules HLA de classe I de haute resolution, et plus recemment de cristaux de molecules HLA de classe 11, permet de localisertres precisement les regions, les acides amines et leurs chames laterales responsables des caracteristiques serologiques et fonctionnelles des molecules HLA.
ORGANISATION GENETIQUE: LA REGION HLA
Definition de la region HLA
Region genetique qui s’oppose aux transplantations de tissus entre des individus de la meme espece, la region HLA (region H2 chez la souris) a ete initialement caracterisee par des methodes d’immunogénétique classique, serologiques et cellulaires. Deux grands ensembles ont ete definis: les antigenes HLA de classe I (HLA-A, B et C), responsables du rejet des allogreffes, et les antigenes HLA de classe 11, responsables des reactions Iymphocytaires mixtes (MLR) et du controle des reponses immunitaires a certains antigenes. Les recombinants naturels familiaux (chez l’homme) et les animaux recombinants (lignees congeniques) ont permis d’etablir les limites respectives de ces ensembles et leur organisation genetique, tres differente chez la souris, ou la region I (classe 11) est inseree entre les principaux genes de classe I (K et D, L), et chez l’homme, ou la region HLA-D (classe 11) est centromerique par rapport a la region des genes de classe 1. Les approches biochimiques ont ensuite revele les caracteristiques structurales des molecules HLA de classe I et 11, produits des genes respectifs qui ont ensuite ete clones, localises et sequences. Le sequen,cage de nombreux alleles des memes genes a permis d’etablir les bases structurales du polymorphisme.
Polymorphisme HLA
Une des caracteristiques du systeme HLA est le nombre d’alleles que de nombreux loci peuvent exprimer. Ce polymorphisme est illustre depuis le premier antigene Mac decrit en 1958 par J. Dausset jusqu’à la nomenclature la plus recense (nomenclature HLA 1992). Son recensement est encore incomplet. Mis en evidence « race a des donnees serologiques et cellulaires, le polymorphisme est desormais etabli par l’étude de la variabilite de la sequence en acides amines (AA) (deduite de la sequence nucleotidique) des domaines des molecules HLA qui permet de caracteriser les nouveaux alleles et les regions les plus polymorphes des molecules HLA de classe I et 11. Certains alleles d’un locus sont associes preferentiellement avec des alleles specifiques d’un autre locus: liaisons entre les alleles de classe I et 11 (Al B8),entre des alleles de la region de classe 11 (DQA et DQB) ou entre classe I et classe 11 (B8-DR3). Ce phenomene appele << desequilibre de liaison >> reste inexplique et varie ainsi que la frequence des differents alleles selon et dans les differentes ethnies.
Carte physique
Le clonage de la region HLA dans des cosmides contigus et chevauchants de 30 a SO Kb chacun et l’analyse de leur contenu ont ete recemment completes par l’obtention de l’integralite de la region HLA sous forme d’une banque de YAC (,veasr artificial chromosome), dont la sequence complete ne saurait tarder. La carte physique actuelle de la region HLA est representee sur la figure 81O-I. Sur le bras court du chromosome 6 (region 6 p 21.3) sont situees:
la region de classe I, d’environ 2 OOO Kb, telomerique codant pour les chaires lourdes des molecules HLA de classe I (A- B-C) ainsi que pour d’autres genes (E. F, G et H) HLA classe I recemment decouverts;
la region de classe II, d’environ I OOO Kb, centromerique codant pour les chaines A et B des molecules HLA de classe 11 et pour les nouveaux genes TAP (TAPI-TAP2) et LMP (LMP2 et LMP7) impliques dans la maturation et le trafic intracellulaire des molecules HLA de classe 1;
ces deux regions sont separees par la region de classe 111 (SW a I OOO Kb), codant pour des composants du complement (C4 B7 C2) et de la 21 hydroxylase (21 OH). De nombreux autres genes, dont ceux du TNF et de la proteine de choc thermique Hsp 70, sont situes entre les regions de classe 111 et 1. Les genes HLA sont composes de sequences codantes (exons) separees par des regions non codantes (introns). L’organisation genomique en exons reflete celle de la structure proteique en domaines.

Figure 810-1 Carte des gènes du complexe majeur d’histocompatibilité de l’homme. (D’après RD Campbell et J Trowsdale. Map of the human major histocompatibility complex, Immunol Today, 1993, 14 : 350-351, centerfold.)
GENES ET MOLÉCULES HLA DE CLASSE I
HLA: A, B, C
La structure des AG HLA de classes I et 11 est representee dans la figure 81()-2. Ces antigenes transmembranaires sont composes: d’une chame glycoproteique de 4S kilodaltons (kDa) divisee en trois domaines extracellulaires correspondant aux 3 exons des genes de classe 1, d’une region transmembranaire et une region intracytoplasmique codees par 2 ou 3 exons et portant un site de phosphorylation. Le polymorphisme est limite a une ou plusieurs zones d’hypervariabilite dont la principale reside au niveau du domaine le plus externe. Cette chaine lourde glycosylee s’associe de maniere non covalente avec la ß2-microglobuline qui, elle, est monomorphe et codee par le chromosome IS. Le niveau d’expression de la ß2- microglobuline contr61e en partie celui de la chame lourde a la surface cellulaire, expliquant son accumulation intracytoplasmique dans les cellules qui ont un defaut de synthese de la ß2-microglobuline. L’association entre chame lourde et microglobuline ß2m a lieu tres precocement dans le reticulum endoplasmique. La structure tridimensionnelle de la partie extracytoplasmique des molecules de classe I est connue en detail. L’unite fonctionnelle composee des domaines al et a2 organises symetriquement forme une cavite, dont le fond est ferme par l’alignement antiparallele des feuillets plisses ß et les bords par deux structures a helicoidales. Le sequen,cage des genes de classe I associe au modele 3 D permet de localiser chaque AA (ou ensemble d’AA polymorphes) au niveau des helices a et des feuillets ß et d’etablir les bases structurales precises du polymorphisme. La nomenclature HLA 1992 denombre 41 alleles A, 61 alleles B et 18 alleles C. La variabilite est concentree au niveau des exons codant pour les domaines al a2 constitutifs du site de liaison du peptide et de la region de contact avec le TCR (CD3). Cette region delimite une cavite destinee a accueillir le (ou les) peptide(s) antigenique(s) d’origine cytosolique et charges dans cette cavite au niveau du reticulum endoplasmique. De la meme maniere, la ß2- microglobuline et le(s) peptide(s) participent au transport de la molecule HLA de classe I et a son expression a la surface cellulaire. Le passage transmembranaire des peptides cytosoliques vers le reticulum endoplasmique est assure en partie ou en totalise par des pompes dont les genes denommes TAPI et TAP2 sont codes par la region HLA de classe 11. Ils forment un canal transmembranaire qui permet un transfert plus ou moins selectif des peptides cytosoliques (peptides du soi) aux molecules HLA de classe 1.
HLA l: E, F, G, H
Des etudes au niveau genomique ont permis de detecter de nouveaux genes classe I appeles HLA-E, F. G, H et J (ou X). Le gene HLA-E est localise entre HLA-A et HLA-C. Des transcrits de ce gene sont trouves dans une grande variete de tissus et leur niveau d’expression, comme pour les genes HLA classe I classiques, peut etre augmente par traitement par l’interferon gamma. Mais l’expression stable d’une molecule HLA-E a la surface des cellules n’a pas ete clairement prouvee. Le gene HLA-E est le plus divergent au sein des genes HLA classe 1. Son polymorphisme est restreint, avec seulement 4 alleles decrits. Le gene HLA-F est le plus telomerique des genes HLA actuellement connus. La distribution tissulaire des transcrits semble recuite aux cellules T au repos et activees ainsi qu’aux cellules EBV. Mais l’expression membranaire de la molecule est peu probable. Le gene HLA-G, situe entre les genes HLA-F et HLA-A, est non polymorphique et code pour la chaine a de la molecule HLA-G exprimee a la surface des cellules cytotrophoblastiques au cours du premier trimestre de la grossesse. Cela suggere que les molecules HLA-G pourraient jouer un r61e protecteur contre les rejets et infections du placenta. Les genes HLA-H et J (ou X) sont des pseudogenes situes a proximite de HLA-A.
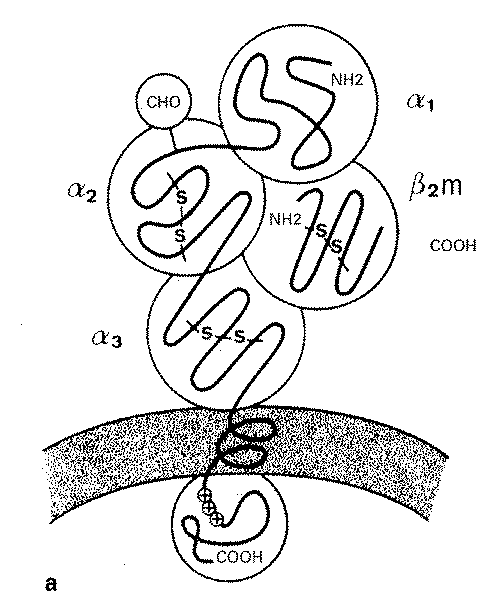
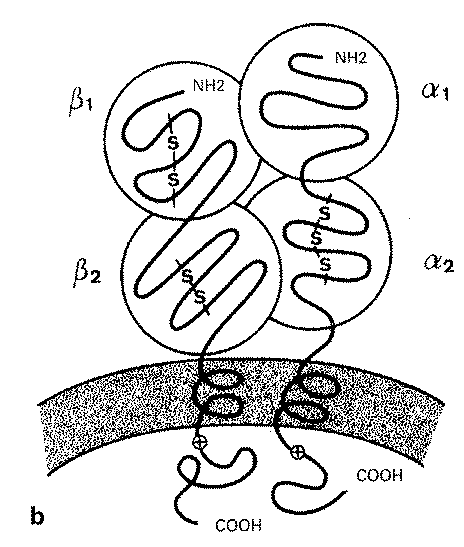
Figure 810-2 Structure des AG HLA de classe I (a) et de classe II (b). (D’ apres A Carpentier et D Farge, Transplantation d’ organes, 1 992, Paris, Medecine-Sciences Flammarion.)
GENES ET MOLECULES HLA DE CLASSE II
Les antigenes HLA de classe II sont des structures heterodimeriques comportant une chaine a et une chaine ß associees de maniere non covalente (voir figure81O-2). Toutes deux sont des glycoproteines transmembranaires de34 kDa et 28 kDa respectivement, qui s’associent pour former 3 isotypes principaux DRa-DRb, DQa-DQb, DPa-DPb.
La sous-region DR comporte un gene A monomorphe codant pour la chaine a des molecules HLA-DR et un ou plusieurs loci B en fonction des haplotypes. Ce polymorphisme haplotypique se manifeste par une organisation differente et un nombre variable de genes DRB sur un chromosome. Cinq groupes haplotypiques ont ete identifies. En plus de ce polymorphisme haplotypique, il existe un important polymorphisme allelique. L’ensemble des alleles actuellement connus est fixe par la nomenclature HLA (tableau 810-1) avec 62 alleles HLA-DRBI, 4 alleles DRB3, 1 allele DRB4, 4alleles DRBS, 14 alleles DQAI, 19 alleles DQBI, 8 alleles DPAI et 37 alleles DPBI. Comme pour les genes HLA classe I, le polymorphisme est concentre dans trois regions hypervariables qui sont pour les genes DR les nucleotides codant pour les AA en position 9-13, 26-33 et 67-74. Caracteristique deja signalee du systeme HLA, le desequilibre de liaison est marque entre les alleles DRB, DQA et DQB. En revanche, il existe tres peu de desequilibre de liaison entre les genes DR, DQ et DP. Les desequilibres de liaison ont ete bien etudies dans la population caucasoide et certaines associations alleliques sont specifiques d’une population. L’existence d’un desequilibre de liaison entraine une reduction du polymorphisme HLA et augmente dans une population donnee les chances de trouver des donneurs de moelle ou d’organes compatibles avec un receveur non apparente. II existe un polymorphisme supplementaire issu de l’association des chaires a et p des 2 haplotypes par transcomplementation, a l’ interieur d’ un isotype (DQ, voire DP). Ces molecules hybrides, decrites chez des sujets heterozygotes, sont generatrices d’un polymorphisme fonctionnel et sont le support moleculaire de nombreuses associations HLA et maladies.
TECHNIQUES D’ETUDE DES ANTIGENES HLA
Typage serologique
La serologie a ete la base de la definition du systeme HLA et est encore utilisee dans la plupart des laboratoires. La serologie classique peut reveler de nouvelles specificites qui seront affirmees par les techniques de sequen,cage et aider a identifier des epitopes fonctionnels. Les anticorps monoclonaux contribuent a la serologie et certains sont deja utilises en routine. Des cellules transvectees et des animaux transgeniques sont utilisables pour rechercher et mieux caracteriser des serums, les anticorps obtenus par ces approches revelant de nouvelles specificites.
Les techniques serologiques, appliquees a l’etude du CMH, reposent sur l’aptitude des anticorps a reconnaitre des determinants allotypiques de ces molecules, a la surface cellulaire. La technique la plus utilisee est la microlymphocytotoxicite dependante du complement (Terasaki et McClelland, 1964). Le principe est le suivant: lorsqu’un anticorps reconnais un determinant antigenique a la membrane d’une cellule, denommee cible, la sequence du complement est activee au site de la reaction antigene/anticorps et aboutit a la Iyse de la cible.
La principale source d’anticorps anti-HLA provient de l’immunisation facto-maternelle, moins souvent de l’immunisation apres transfusion ou transplantation. Les ACMC (anticorps monoclonaux) sont de plus en plus utilises car specifiques des produits du CMH. Cette analyse serologique fine n’est possible que si le laboratoire possede un paner de reference, etudie avec des anticorps definis au cours des ateliers internationaux afin de contr61er les reactifs locaux par rapport a ceux definis internationalement.
Les techniques usuelles sont utilisees pour detecter les alleles de classe II. Les antigenes de classe II n’etant pas ubiquitaires, la microlymphocytotoxicite est realisee sur Iymphocytes B prealablement isoles.
Le crossmatch permet de rechercher par un test de Iymphocytotoxicite ou par cytometrie de flux la presence chez le receveur d’anticorps (notamment de type IgG) specifiques du donneur et susceptibles d’entrainer un rejet suraigu. La specificite anti-HLA classe I (testee sur cellules B et T) de ces anticorps est une contre-indication a la greffe, alors qu’une specificite anticlasse II (testee sur Iymphocytes B uniquement) serait moins deletere.
Typage moleculaire
Les avantages decisifs des techniques de biologie moleculaire (etude directe des genes polymorphes avec une precision absolue, absence de limite a l’obtention de reactifs) ne sont encore applicables en routine qu’aux alleles HLA de classe II et le seront bientôt aux alleles HLA de classe I
Les techniques faisant appel a la biologie moleculaire reposent sur la connaissance du polymorphisme des sequences nucleotidiques de l’ADN des genes HLA. L ‘ amplification specifique d ‘ un fragment d’ ADN genomique est obtenue par reaction en chatne de la polymerase (PCR). Les alleles sont detectes a l’aide de sondes oligonucleotidiques specifiques (dot blot ou reverse dot blot) PCR-SSO ou d’enzymes de restriction PCR- RFLP pour HLA-DRB. La finesse du typage (typage generique 18 alleles versus typage specifique 45 alleles) depend de l’indication meme de celui-ci. Plus de IOO alleles sont definis au niveau des loci HLA classe II, mais certains d’entre eux sont moins frequents chez les Caucasiens.
L’etablissement d’une strategie de typage de routine depend de son but. typage de donneurs volontaires de moelle en vue de constituer un fichier national (typage generique automatise); recherche d’identité entre donneur et receveur en cas de greffe de moelle osseuse (typage complet) ou d’organes (typage generique); recherche d’une predisposition genetique dans le cadred’associations HLA et maladies (typage cible); etudes anthropologiques; medecine legale.
ASPECTS CLINIQUES
Greffes de rein
L’importance de la compatibilite HLA dans le devenir des allogreffes renales est bien etablie. La survie a IO ans est d’environ 75 p. IOO chez les germains (freres et sceurs) HLA identiques et de plus de SO p. IOO si un seul haplotype est partage, comparee a une survie a lOans de 4Op.lOO pour les reins de cadavres. L’introduction de la ciclosporine a ameliore ces resultats, qui restent neanmoins meilleurs selon le degre de compatibilite. Les series alleliques A B ainsi que DR DQ sont impliquees dans ce phenomene.
Si la precision obtenue par le typage moleculaire va permettre une appreciation plus juste du r61e respectif des loci et alleles du CMH, la survenue d’episodes de rejet chez les germains HLA identiques reflete l’existence d’antigenes mineurs d’histocompatibilite. L’etablissement futur d’une hierarchie des incompatibilites devrait permettre de selectionner les moins immunogenes sur le plan fonctionnel.
Autres transplantations d’organes
Le r61e benefique de la compatibilite HLA est demontre pour les transplantations de c_ur de pancreas et de cornee, meme si cette compatibilite n’entre pas encore dans les criteres de selection pour des raisons logistiques. Le r61e de la compatibilite HLA reste controverse en transplantation hepatique.
Greffe de moelle
La moelle osseuse contient des cellules souches immunologiquement competentes, myeloides et Iymphoides, qui expliquent la survenue, a côté de la reaction de rejet decrite pour toute greffe d’organe, de reaction du greffon contre l’hôte (GVH). La GVH, due a l’activation des cellules T contenues dans la moelle du donneur et dirigees contre les antigenes majeurs ou mineurs d’histocompatibilite du receveur, comprend plusieurs etapes: reconnaissance des antigenes d’histocompatibilité du receveur par les cellules T du donneur, activation des cellules T du donneur et des cellules cibles du receveur, cytotoxicite cellulaire par l’intermediaire des Iymphokines secretees par les cellules B et T activees. La consequence finale de la GVH est la destruction des tissus cibles (kératinocytes, cellules glandulaires de l’intestin, cellules des canalicules biliaires du foie) avec un deficit immunitaire profond, responsable d infections parfois letales. Le risque de rejet augmente en cas de disparite HLA entre donneur et receveur. Un crossmatch serologique positif contre les cellules T ou B du donneur semble correle avec le rejet dans les greffes HLA incompatibles.
La GVH, letale dans 2O p. IOO des cas, explique les criteres de choix du donneur.
Le premier choix se portera sur une greffe syngenique a partir d’un jumeau monozygote. L’homozygotie peut etre affirmee par les methodes serologiques classiques, les techniques de culture mixte Iymphocytaire et de typage par biologie moleculaire. Lefingerprinting et le DNA crossmatch utilisant la technique des heteroduplexes de l’ADN, I’etude des minisatellites d’ADN permettent d’affirmer l’identite totale entre deux individus et semblent les methodes de choix pour affirmer la monozygotie. La greffe syngenique apparais ideale puisqu’elle ne s’accompagne ni de rejet ni de GVH. Par contre, dans les cas d’aplasie, certaines non-prises plaident pour l’origine auto-immune de la maladie. Ainsi, actuellement, entre un frere HLA genotypiquement identique et un jumeau, le choix pourra se porter sur le frere genotypiquement identique.
Chez l’animal comme chez l’homme, le donneur HLA genotypiquement identique est le meilleur possible, maigre la survenue de rejet et de GVH dans SO p. IOO des cas, responsables d’une mortalite de 20 p. IOO. Cela prouve qu’a côté du complexe majeur d’histocompatibilite existent d’autres systemes mineurs d’histocompatibilité qui ont un r61e important apres greffe de moelle allogenique.
Un donneurfamilial HLA phenotypiquement identique peut etre trouve parmi les freres et s_urs du patient, si les parents ont un haplotype commun, le plus souvent en cas de mariage consanguin. Les resultats sont equivalents a ceux des greffes apparentees genotypiquement HLA identiques.
Vu la taille des familles en France, seuls 3O p. IOO des patients pourront trouver a l’interieur de leur famille un donneur geno-identique et IO p. IOO un donneur pheno-identique. Pour les 6() p. IOO restants, des 1986 s’est cree en France un registre de donneurs volontaires pour trouver un donneur HLA phenotypiquement identique non apparente, registre connecte en 1992 a un fichier europeen.
Des greffes avec donneur apparente non HLA identique ont ete realisees a partir de donneurs intrafamiliaux avec un haplotype HLA identique alors qu’existait une incompatibilite sur un, deux ou trois antigenes (A, B, DR) sur l’autre haplotype. Dans les deficits immunitaires severes de l’enfant, I’augmentation du conditionnement, la depletion de la moelle en cellules T et l’utilisation d’anticorps monoclonal contre la molecule d’adhesion LFA-I donnent des resultats satisfaisants. Ces resultats ne sont pas observes dans les autres indications, I’incidence du rejet et de la GVH augmentant avec le nombre d’incompatibilités.
PERSPECTIVES
Les progres rapides de nos connaissances sur le systeme HLA et son role en transplantation autorisent a predire, en ce qui concerne les typages HLA, la disparition progressive de la serologie en raison de sa complexite, de sa fiabilite et de sa reproductibilite relatives et de son coût au profit de la biologie moleculaire fiable, reproductible, automatisable. La serologie a l’aide d’anticorps monoclonaux, dont les reactifs sont specifiques et standardises, conservera une place dans les typages d’urgence. Le r61e des differents alleles et loci sera precise afin d’etablir la hierarchie des incompatibilites et de permettre un meilleur choix du couple donneur-receveur en augmentant le nombre de greffes compatibles. Outre les classiques antigenes HLA A,B,C et DR DQ DP, le systeme HLA comporte de nombreux autres genes (TAP, LMP, TNF…) impliques dans le contr61e des reponses immunes, dont les poids respectifs dans la stimulation allogenique constitueront un veritable index << immunogenetique >>. La connaissance de la structure et des interactions moleculaires entre les molecules HLA et leurs partenaires (TCR, pep tides, superantigenes) permettra le developpement de nouvelles methodes immunosuppressives specifiques (analogues de peptides bloquant la reponse immune) ou meme d’inducteurs de tolerance (anticorps monoclonaux).